El ADN portador de la información genética.
Duplicación del ADN.
Síntesis de proteínas.
Características del código genético.
Regulación de la expresión genética.
1. EL ADN PORTADOR DE LA INFORMACIÓN GENÉTICA
La molécula de ADN es la portadora de la información genética (excepto en los virus de ARN). Se llegó a la siguiente conclusión, a partir de las siguientes observaciones y experiencias:
· Presencia de ADN en el núcleo y en los cromosomas.
· La cantidad de ADN en individuos de la misma especie es constante.
· Cuanto más compleja es una especie, mayor cantidad de ADN contiene.
· La luz ultravioleta de 360 nm. es la más absorbida por el ADN y es, precisamente esta radiación, la que provoca las mutaciones.
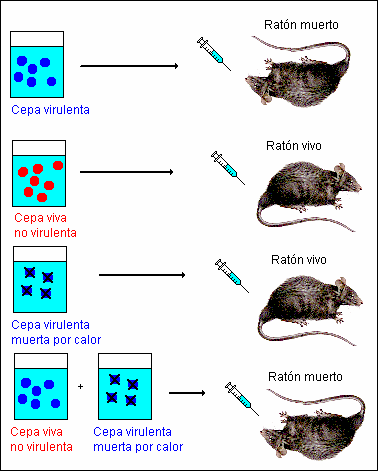
· La prueba definitiva la aportaron las observaciones de Griffith, 1928, sobre transformaciones en bacterias:
2. DUPLICACIÓN DEL ADN
Este proceso permite el paso de información genética a las células hijas. A partir de la estructura de la doble hélice propuesta por Watson y Crick, es fácil pensar cómo puede producirse: Se separan las dos hebras y los nucleótidos se van uniendo por complementariedad.
Se propusieron tres hipótesis: Conservativa, semiconservativa y dispersiva.
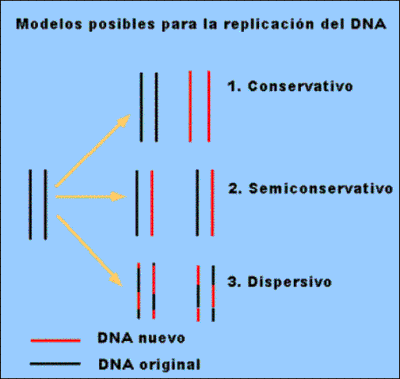
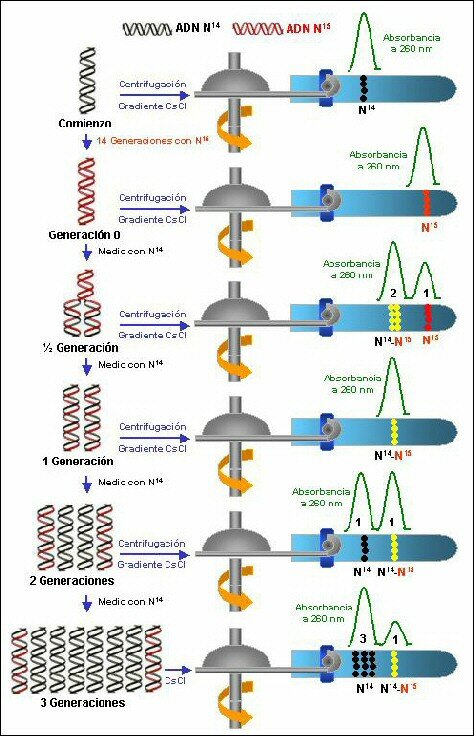
La comprobación experimental fue realizada por Meselson y Stahl:
La síntesis del ADN requiere de: la enzima ADN polimerasa III (ADNpol III); la presencia de desoxirribonucleótidos 5’ trifosfatos de A, T, G y C; iones Mg++; un ADN patrón; y un extremo 3’ libre de otra cadena que actúa como cebador (la ADNpol III es incapaz de iniciar una cadena y se limita a ańadir desoxirribonucleótidos a una cadena existente).
La dirección de crecimiento de la hebra o cadena será siempre 5’ ![]() 3’, es decir sólo se unen los dnucleótidos al extremo 3’ de la cadena de ADN ya formada.
3’, es decir sólo se unen los dnucleótidos al extremo 3’ de la cadena de ADN ya formada.
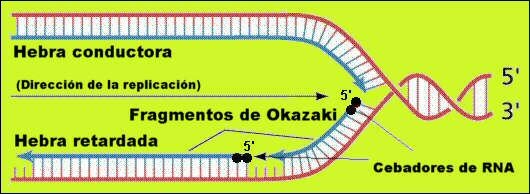
En un principio se pensó que el crecimiento era unidireccional, cuando en realidad es bidireccional. Pero żCómo podía sintetizarse la cadena de 3’![]() 5’ si ninguna ADNpol lo hacía? La respuesta la dio el científico Okazaki, que propuso el siguiente mecanismo para la duplicación:
5’ si ninguna ADNpol lo hacía? La respuesta la dio el científico Okazaki, que propuso el siguiente mecanismo para la duplicación:
En bacterias
1. Existe una secuencia de ADN que será el origen de la duplicación, denominada seńal de iniciación.
2. La enzima helicasa separa las dos hebras, la girasa elimina las tensiones y las proteínas SSB estabilizan la separación de las dos cadenas.
3. Esto da origen a la burbuja de replicación.
4. El fragmento cebador lo sintetiza la ARNpol, y es un fragmento de ARN de unos 10 nucleótidos denominado PRIMER.
5. Ahora la ADNpol III ya puede ir ańadiendo los nucleótidos al extremo 3’ formándose la denominada cadena continua o conductora. La energía para el proceso se obtiene de los propios dnucleótidos 5’ trifosfatos.
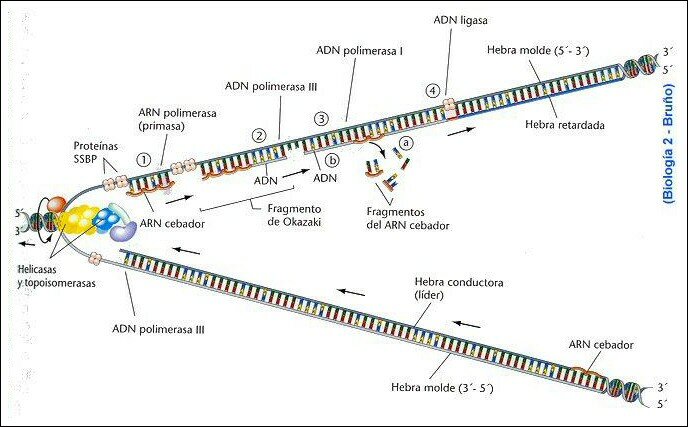
6. Sobre la otra hebra la ARNpol debe esperar a que la burbuja tenga un tamańo de unos 1000 nucleótidos a cada lado para poder sintetizar el primer, al que ańadirá dnucleótidos la ADNpol III, formándose el denominado fragmento de Okazaki. Cada vez que se abre la cadena, se repite el proceso formándose la denominada cadena discontinua o retardada.
7. Para completar las cadenas, debe actuar la ADNpol I que es capaz de retirar todos los fragmentos de ARN y sustituirlos por ADN y finalmente interviene la ADNligasa para unir todos los fragmentos separados.
En Eucariotas
El proceso es similar con ciertas particularidades:
- Las histonas del ADN original se quedan en la hebra conductora, sintetizándose nuevas para la retardada.
- Existen muchos puntos de origen de la replicación, lo que da origen a las unidades de replicación o replicones.
- Los fragmentos de Okazaki tienen un tamańo de sólo 100 a 200 nucleótidos.
3. SÍNTESIS DE PROTEÍNAS
La información del ADN podía generar directamente una proteína, sin embargo debía existir un intermediario, ya que el ADN se encuentra en el núcleo, mientras que los ribosomas están en el citoplasma. Este era el ARNm y el proceso se realiza en dos etapas:
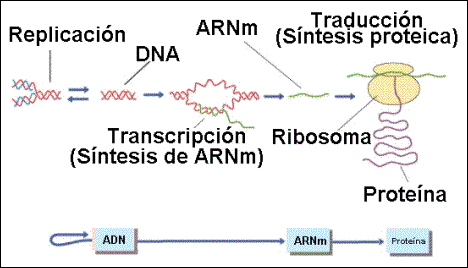
Transcripción
Consiste en la síntesis de moléculas de ARN complementarias de ADN. Las enzimas que lo llevan a cabo son las ARNpol (son muy complejas, formadas por muchos polipéptidos). La ARNpol I fabrica los ARNr, la ARNpol II fabrica los ARNm y la ARNpol III fabrica los ARNt.
Las proteínas se obtienen a partir de los ARNm y en su formación tienen lugar los siguientes procesos:
1. Iniciación: La ARNpol reconoce y se une a una secuencia específica llamada promotor.
2. Alargamiento: Se produce por la unión de sucesivos nucleótidos complementarios de las bases de una de las cadenas del ADN. La dirección de crecimiento es 5’![]() 3’.
3’.
3. Terminación: Se produce cuando se llega a la región terminadora del gen.
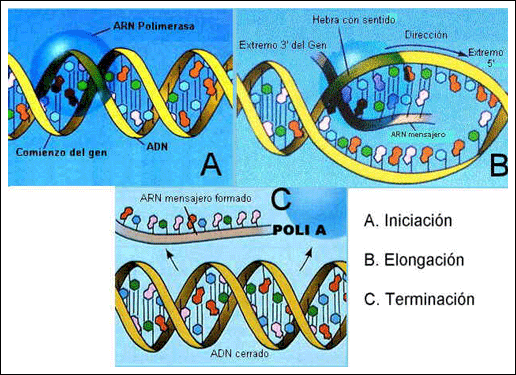
Con estos procesos, el ARN de procariotas ya está listo para ser traducido en los ribosomas, pero no así el de las células eucariotas que debe sufrir una serie de transformaciones de constituyen la maduración y que le permiten salir del núcleo.
Primero en el 5’ se le ańade un 7 metil Guanosina 3P, después se le ańade una cola de poliAdenina de unos 200 nucleótidos en el extremo 3’ y por último se eliminan los intrones, o zonas sin información para la proteína.
Ahora ya se puede denominar ARNm y salir del núcleo a través de los poros celulares.
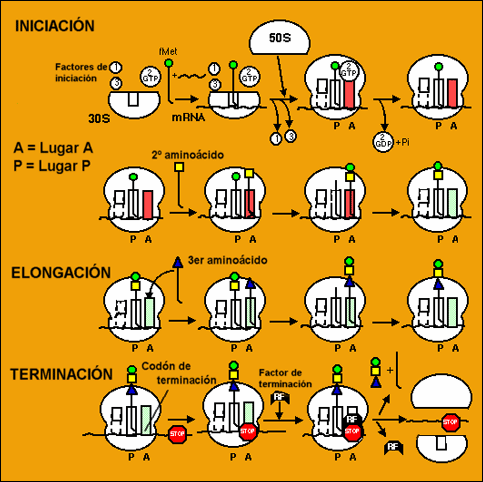
Traducción
Consiste en la fabricación de la proteína según la información del ARNm. Los aminoácidos son transportados por los ARNt y el proceso tiene lugar en los ribosomas.
Se distinguen las siguientes etapas:
1.- Activación de los aminoácidos.
Los aminoácidos en presencia de la enzima aminoacil-ARNt-sintetasa y de ATP, son capaces de unirse a un ARNt específico y dar lugar a un aminoacil-ARNt liberándose AMP y 2Pi y quedando libre el enzima.
2.- Iniciación de la síntesis.
El ARNm se une a la subunidad menor del ribosoma. A estos se asocia el aminoacil-ARNt, gracias a que el ARNt tiene en una de sus asas un triplete de nucleótidos llamado anticodón, que se asocia al primer triplete del ARNm según la complementariedad de sus bases. A este grupo de moléculas se une la subunidad ribosómica mayor formándose el complejo ribosomal o complejo activo. Estos procesos están catalizados por los llamados factores de iniciación (FI). El primer triplete o codón que se traduce en eucariotas es el AUG que corresponde al aminoácido Metionina, en procariotas corresponde a la Fenilmetionina.
3.- Alargamiento de la cadena polipeptídica
El complejo ribosomal posee dos centros: el centro peptidil o centro P, donde se sitúa el primer aminoacil-ARNt y el centro aceptor de nuevos aminoacil-ARNt o centro A, El radical COOH del aminoácido iniciador, se une con el radical amino NH2 del aminoácido siguiente mediante enlace peptídico. El centro P queda pues ocupado por un ARNt sin aminoácido y sale del ribosoma.
Se produce entonces la translocación ribosomal en el sentido 5’![]() 3’ El dipeptidil-ARNt queda ahora en el centro P. Todo esto es catalizado por los factores de elongación (FE) y precisa GTP,
3’ El dipeptidil-ARNt queda ahora en el centro P. Todo esto es catalizado por los factores de elongación (FE) y precisa GTP,
Según la información del tercer codón aparece el tercer aminoacil-ARNt que ocupa el centro A. Se forma el tripéptido en A y posteriormente el ribosoma, realiza su segunda translocación.
4.- Terminación de la síntesis.
El final de la síntesis viene informado por los llamados tripletes sin sentido: UAA, UAG y UGA No existe ningún ARNt cuyo anticodón sea complementario de ellos y por tanto la biosíntesis se interrumpe. Indica que la cadena polipeptídica ha terminado. Este proceso viene regulado por los factores R.
Un mismo ARNm puede ser leído (traducido) por varios ribosomas a la vez, uno detrás de otro (polirribosomas).
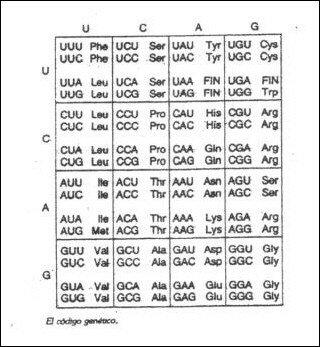
4. CARACTERÍSTICASDEL CÓDIGO GENÉTICO
Se sabe que los componentes de las proteínas son 20 aminoácidos. Como tenemos 20 aminoácidos diferentes, como mínimo necesitamos que la información se cada aminoácido sea por tres bases nitrogenadas, denominadas tripletes o codones. Como al combinar las tres bases nos dan 64 posibles codones, todos los aminoácidos tienen varios tripletes, excepto la Metionina y el Triptófano. Debido a esto se dice que el código genético es degenerado.
El ser degenerado es una ventaja, ya que los errores producidos, por ejemplo por mutaciones, muchas veces no se aprecian.
El código genético es universal, excepto para el ADN mitocondrial, donde hay algunos cambios.
El código genético no es ambiguo, sino determinado, ya que a cada triplete le corresponde siempre el mismo aminoácido.
En el descubrimiento del código genético tuvo una importancia trascendental el aislamiento de la enzima polinucleótido fosforilasa, por Severo Ochoa en 1955. Este enzima permite unir nucleótidos entre sí “in vitro”.
5. REGULACIÓN DE LA EXPRESIÓN GÉNICA
La cantidad de enzimas presentes en una célula, depende del proceso que se esté realizando. Cuando varíe el proceso, los enzimas deben desaparecer y formarse otros nuevos para intervenir en el nuevo proceso. De esta forma se consigue una gran economía que impera en todas las células y además se evita un posible caos metabólico.
El control de los enzimas que deban fabricarse se lleva acabo a nivel del ADN, controlando el ARNm que se vaya a transcribir.
En BACTERIAS se descubrió el modelo de OPERÓN que regula el uso de la glucosa o lactosa como sustrato por parte de la bacteria E. coli.
En el cromosoma de E. coli se distinguen las siguientes partes relacionadas con el uso de la lactosa:
Cuando no hay lactosa, el represor, que es una proteína sintetizada a partir del gen regulador, se une al operador e impide la transcripción de los genes estructurales.
En presencia de lactosa, ésta se une al represor y lo anula, de forma que los genes estructurales pueden sintetizarse.
Además se descubrió otro control a través del AMPc. Este se forma a partir de ATP por la enzima Adenilato ciclasa, que se encuentra en la cara interna de la membrana.
El AMPc se une a la proteína activadora del catabolismo (CAP), formándose es complejo [AMPc-CAP], que se une al promotor y ayuda a la unión de la ARNpol que transcribirá los enzimas estructurales.
Cuando aumenta la concentración de glucosa, disminuye la concentración de AMPc y no se podrá unir la ARNpol y no se transcribirán los genes estructurales. Cuando no hay glucosa ocurrirá lo contrario.
En EUCARIOTAS
Las células de los organismos pluricelulares responden a las variaciones hormonales del medio. Todas las células llevan la misma información genética, pero no se manifiestan todos sus genes. La diferenciación celular, aparece porque se condensan distintas zonas de los cromosomas en las diferentes células, que además presentarán distintos receptores en sus membranas.
El control de la expresión es distinto según el tipo de hormona:
a) Hormonas lipídicas: Atraviesan la membrana y se unen a proteínas receptoras formándose el complejo H-R que va al núcleo y se une a secuencias determinadas del ADN, induciendo la transcripción de determinados genes. Por ejemplo, las hormonas anabolizantes provocan la síntesis de proteínas musculares.
b) Hormonas proteicas: No pueden atravesar la membrana. Se unen a recetores formándose el complejo H-R, que provoca la reacción en el interior por parte de la Adenilato ciclasa del paso de ATP a AMPc. El AMPc viaja al núcleo y activa las proteínas reguladoras de la transcripción.